Pour diversifier un peu mes observations, j'ai acheté un petit coffret de lames préparées. Ce coffret contient 30 lames de divers tissus histologiques humains. Déjà fixées et colorées, ces préparations sont toutes prêtes à être observées. On ne peut pas faire plus simple... Elles sont vendues sous la marque Bresser, qui est une société allemande spécialisée dans la vente de matériel d'optique (astro, ornitho, microscopie...). Bien qu'existant depuis la fin des années 60, cette marque n'est pas très connue en France, et je n'en ai surtout entendu parler que par le biais de leurs jumelles revendues chez Lidl et qui ont eu un certain engouement chez les amateurs. En réalité, Bresser n'est qu'un distributeur de matériel chinois. Entre 1999 à 2009, Bresser a été rachetée par Meade, ça explique qu'au dos de la boîte on retrouve la mention "Meade Instruments Europe" sur l'étiquette. Aujourd'hui Bresser appartient en partie au géant chinois JOC. Bref, Bresser c'est du matériel chinois de moyenne gamme, on verra bien si ces lames valent le coup, ou pas... |
||
 |
||
Je vais tout de suite tester la première lame. L'étiquette indique "Loose Connective Tissue, W.M.", c'est à dire du tissu conjonctif lâche. WM, c'est l'abréviation de "whole mount", qui signifie que le tissu n'a pas été coupé auparavant et qu'il a été monté directement sur la lame. |
||
 |
||
Je la place d'abord sous la loupe binoculaire. Je ne prendrai des photos qu'en lumière transmise car la lumière incidente fait apparaître trop d'artéfacts, notamment des reflets. Au zoom 0,7X, la préparation rentre en entier sur le capteur ! |
||
 |
||
À ce grossissement, on voit un tissu hétérogène, de couleur violacée et de texture filamenteuse à la périphérie, un peu comme une toile d'araignée. Il se densifie vers le centre de la préparation, et y prend une texture plus granuleuse, parcouru de petits points. Je passe au zoom 2X sur la partie centrale. On détaille mieux ces milliers de petits points sombres, répartis irrégulièrement sur un fond rose. |
||
 |
||
Au 2X, la partie périphérique montre un aspect différent. Le fond est blanc, les petits points sombres sont moins abondants et la texture filamenteuse que l'on observait à 0,7X devient un enchevêtrement de minces fils formant une trame. |
||
 |
||
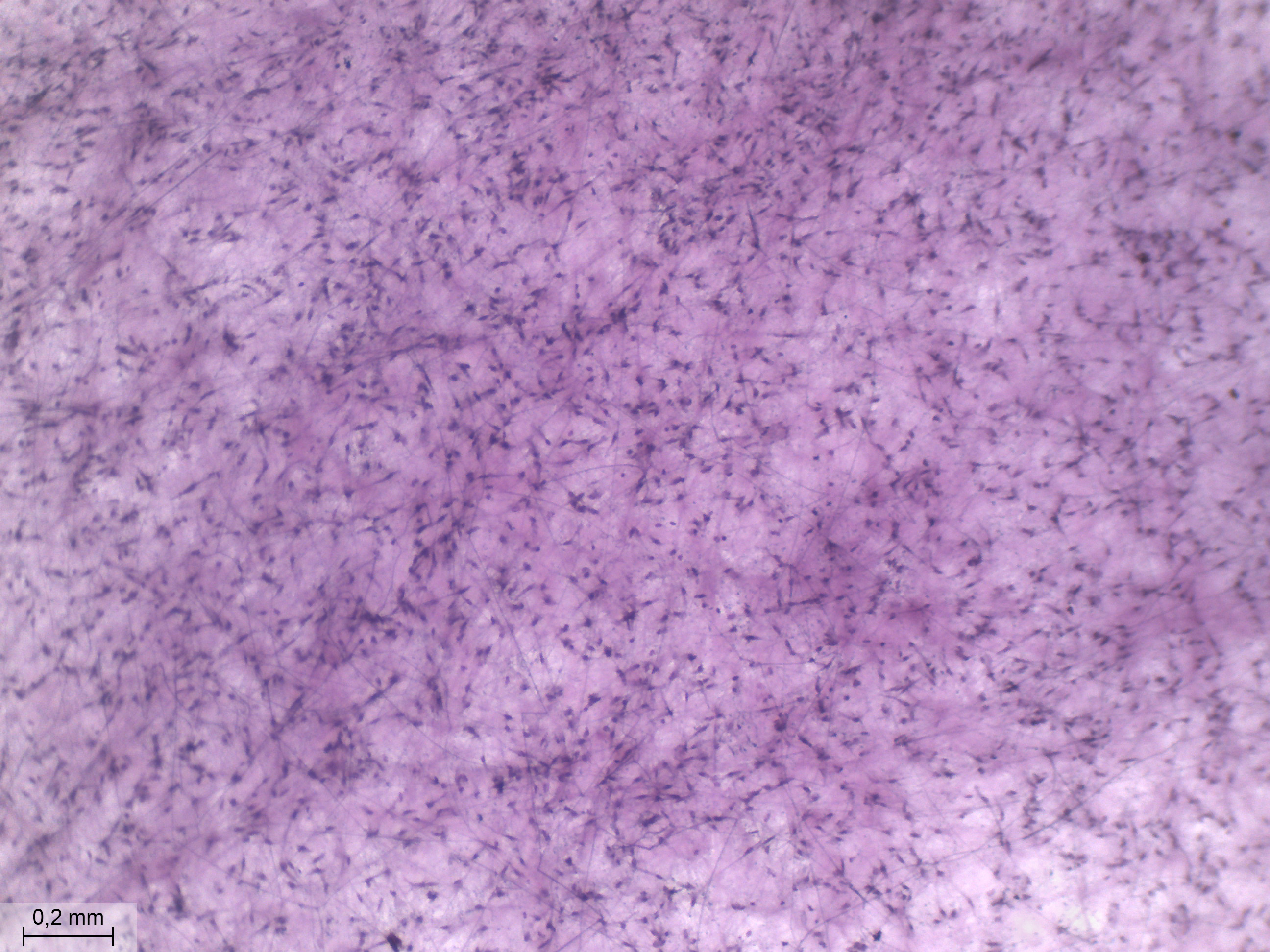
Au zoom maximum (4,5X) sur la partie centrale de la lame, on constate que les points sombres sont des cellules, ou des noyaux de cellules. Dans cette zone aussi, il y a des filaments, mais ils sont peu nombreux et très fins. |
||
 |
||
À ce grossissement sur la partie périphérique, la concentration cellulaire s'est réduit, tandis que la réseau de filament s'est franchement densifié. |
||
 |
||
Bien ! C'est un bon début, qui a déjà aiguisé ma curiosité et il me tarde de passer au microscope pour détailler tout ça. Le temps de ranger la loupe binoculaire et de sortir le microscope, je m'accorde une pause pour relire mes vieux cours d'histologie de première année, qui me permettront certainement de mieux profiter du spectacle. Ah, j'ai trouvé la bonne page... « Le tissu conjonctif lâche est un tissu conjonctif riche en cellules et pauvre en collagène. Il est très répandu dans l’organisme. On en trouve sous les épithéliums de revêtement (derme papillaire, sous-muqueuses du tube digestif, des voies respiratoires, génitales ou urinaires, où il porte le nom de chorion), dans l’adventice des vaisseaux, entre les masses musculaires, dans le parenchyme de nombreux organes pleins (où il est nommé stroma conjonctif). Il a un rôle mécanique de protection et de soutien des tissus et des organes dont il assure la cohésion en comblant les espaces vides. Richement vascularisé, ce tissu conjonctif exerce aussi d'importantes fonctions métaboliques, échanges entre les tissus, trophicité des épithéliums. Et puisqu'il est le de siège de nombreuses cellules libres du système immunitaire, il joue également un rôle majeur dans les réactions inflammatoires, les phénomènes immunitaires et les processus de cicatrisation. On retrouve 3 parties dans la structure du tissu conjonctif lâche : 1) La matrice extra-cellulaire Elle est très abondante, souple, et se compose d'une substance fondamentale contenant des fibres. La substance fondamentale est dite amorphe, c'est à dire qu'elle a une faible densité et une texture plus ou moins gélifiée. Elle ne fixe pas les colorants usuels et par conséquent apparaît transparente en microscopie optique. Elle est homogène, non structurée et occupe un espace important au sein de ce tissu. Elle se constitue d'eau, de sels minéraux, d'électrolytes, et de molécules organiques (protéoglycanes et glycoprotéines). Les fibres se divisent en 3 groupes suivant leur nature : |
||
|
||
2) Les cellules Les cellules du tissu conjonctif sont dispersées au sein de la trame matricielle. Elles sont non jointives. Certaines sont liées à la matrice, d’autres sont libres. a) Cellules reliées au réseau matriciel : |
||
|
||
b) cellules libres : Elles appartiennent presque toutes au système immunitaire. Leur présence est plus inconstante. On pourra retrouver : |
||
|
||
3) Vaisseaux et nerfs Enfin on retrouve presque toujours des vaisseaux (sanguins et lymphatiques) et des nerfs (fibres nerveuses) qui traversent ce tissu. » Voilà, le microscope est prêt et ma mémoire raffraichie. On va essayer de voir tout ça... Première étape, je glisse la lame sous le plus faible objectif du microscope (4X). Je vais pointer les mêmes zones qu'au plus fort grossissement de la loupe, histoire de comparer les rendus. L'image est inversée à 180° par rapport à la loupe, ça ne facilite pas le repérage de la zone précédente, mais le grossissement est faible, je trouverai vite. La différence est évidente, le microscope donne une image plus contrastée et dont la résolution est supérieure en dépit du grossissement qui est un peu inférieur (40X contre 45X). Ci-dessous, la zone centrale : |
||
 |
||
Et là, la zone périphérique : |
||
 |
||
Allez, on grossit un peu, ça fera pas de mal... |
||
 |
||
| Les filaments mycéliens sont clairs. Leur couleur s'assombrit par intermittence. Ils se ramifient par endroits. Le fond de la lame est parsemé de spores (photo au microscope, objectif 4X). | ||
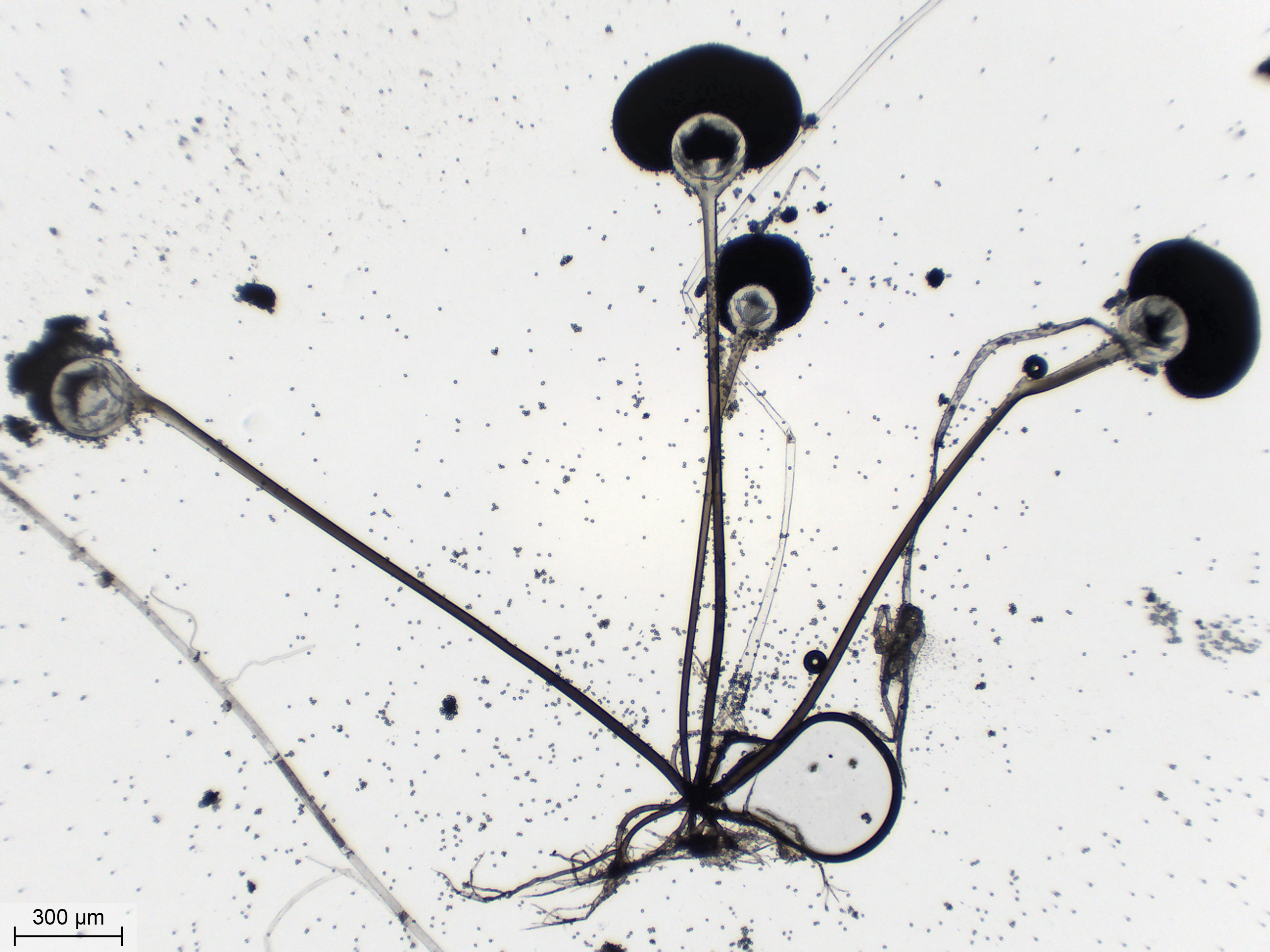
 |
||
| Deux sporocystes à l'objectif 10X. Le fond du champ est parsemé de spores. On voit un stolon passer en arrière, transparent comme un tube de verre. | ||
 |
||
Détail d'un rhizoïde à l'objectif 10X. On voit bien les ramifications qui ressemblent beaucoup aux racines d'une plante. Mais à la différence de racines, le rhizoïde n'a aucun rôle nutritionnel et ne sert qu'à fixer le mycélium au substrat. Quatre sporocystophores, de couleur brune, partent vers le haut. Une 5ème branche, part en haut à droite, sans coloration. C'est un stolon. Il y a une grosse bulle d'air sur la droite. |
||
 |
||
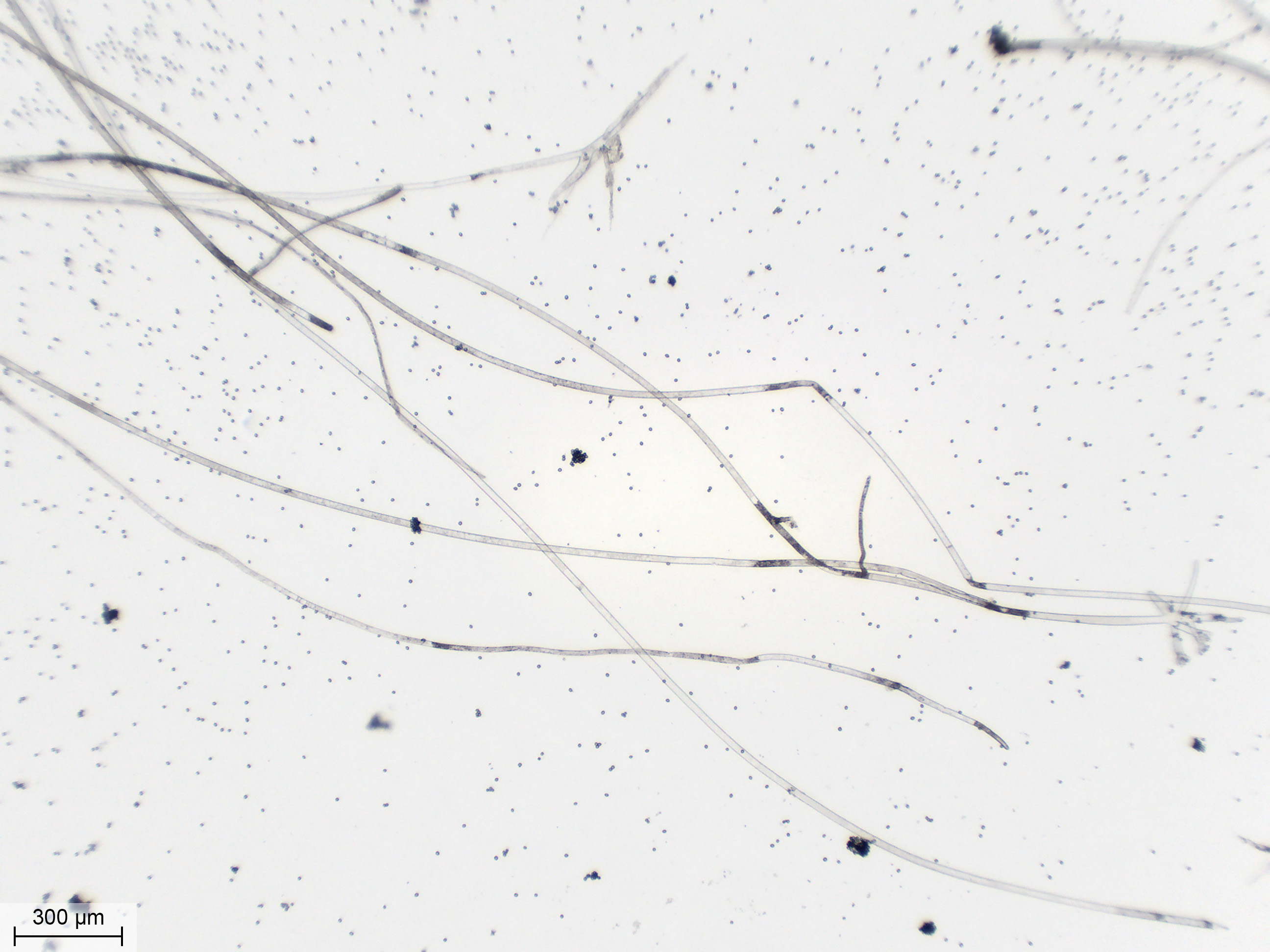
| Quelques filaments mycéliens, toujours sous l'objectif 10X. On voit qu'ils se ramifient. Ils n'ont pas de couleur particulière mais semblent contenir une substance qui est plus foncée par endroits. | ||
 |
||
Je me déplace un peu derrière l'objectif 10X. Ici, un filament mycélien avec des ramifications naissant régulièrement. Son contenu est un peu plus foncé, hétérogène, mais reste toujours bien plus clair que le sporocystophore. À droite, une columelle et quelques restes de spores en train de se détacher par grappes. |
||
 |
||
Détail d'un sporocyste à l'objectif 10X. Le sporocystophore descend de la gauche. On voit bien sa couleur s'éclaircir progressivement. La pigmentation semble due à une substance brune qu'il contient, hétérogène, et qui se raréfie en distalité. Son renflement terminal forme la columelle, sphérique et froissée. Elle contient quelques spores au centre. Tout autour s'agglutinent des centaines de spores, sans membrane périphérique pour les retenir. |
||
 |
||
Dernière photo à l'objectif 10X, avec un pêle-mêle de ce qu'il y a à voir. Plusieurs filaments s'entrelacent. Le plus brun en haut est un sporocystophore. Il se termine par une columelle, vide et fripée. Il y a une autre columelle en dessous. Elle s'est rompue et déverse son contenu, un fluide laiteux et hétérogène qui trouble le milieu aqueux extérieur. D'ailleurs son sporocystophore est beaucoup plus clair, partiellement évidé de la substance qui lui donne sa couleur. Les autres filaments visibles sont plus clairs encore, c'est du mycélium. De nombreux spores apparaissent dans le champ, soit regroupés en amas comme sur la gauche de la photo, soit éparpillés aléatoirement, comme sur la droite. |
||
 |
||
Je passe maintenant à l'objectif 40X du microscope. Zoom sur un filament mycélien. C'est une structure cellulaire tubulaire, possédant une fine membrane plasmique. À l'intérieur, le cytoplasme contient des réserves de glycogène et de lipides bien visibles sur la photo ci-dessous, sous la forme de vacuoles et d'hétérogénéités. C'est cette substance protoplasmique qui trouble le milieu aqueux lorsque la cellule se brise. |
||
 |
||
| Par endroits, le protoplasme est beaucoup plus épais. | ||
 |
||
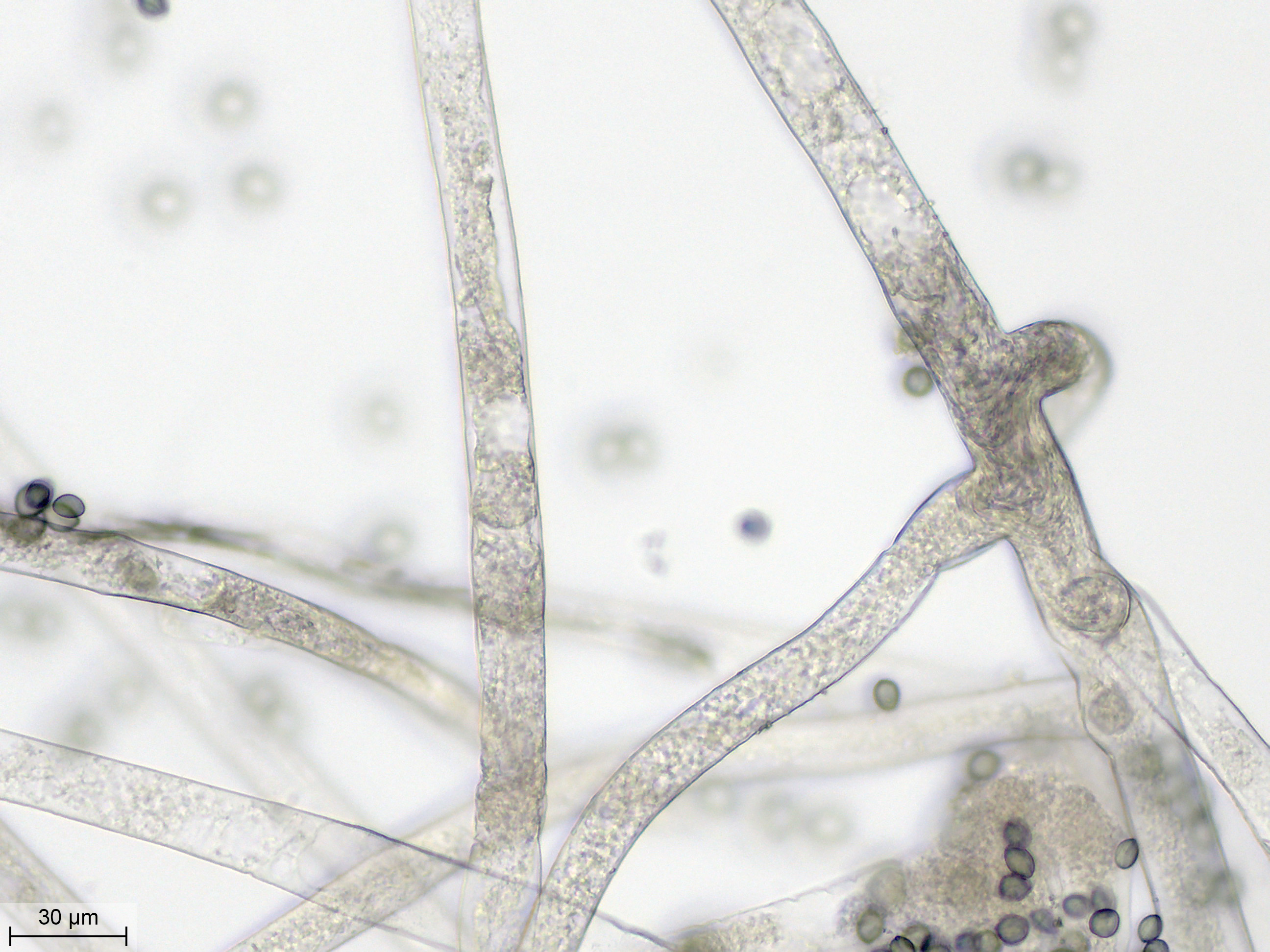
| Les filaments mycéliens mesurent 15 à 20 µm de diamètre. Ils se ramifient. | ||
 |
||
| Le sporocystophore est plus épais, avec un diamètre de 30 µm. Il est finement vacuolisé et malgré sa couleur bien plus foncée, son contenu semble moins hétérogène que celui du stolon. | ||
 |
||
Toujours à l'objectif 40X, je détaille les spores. Ils ressemblent à des petits citrons verdâtres, de 8 à 12 µm de long. Leur paroi externe présente de fines stries longitudinales en relief. |
||
 |
||
En modifiant la focalisation, on peut détailler l'intéreur des spores. Elles contiennent plusieurs inclusions : des vacuoles et des vésicules qui sont des sphérosomes lipidiques et protéiques. |
||
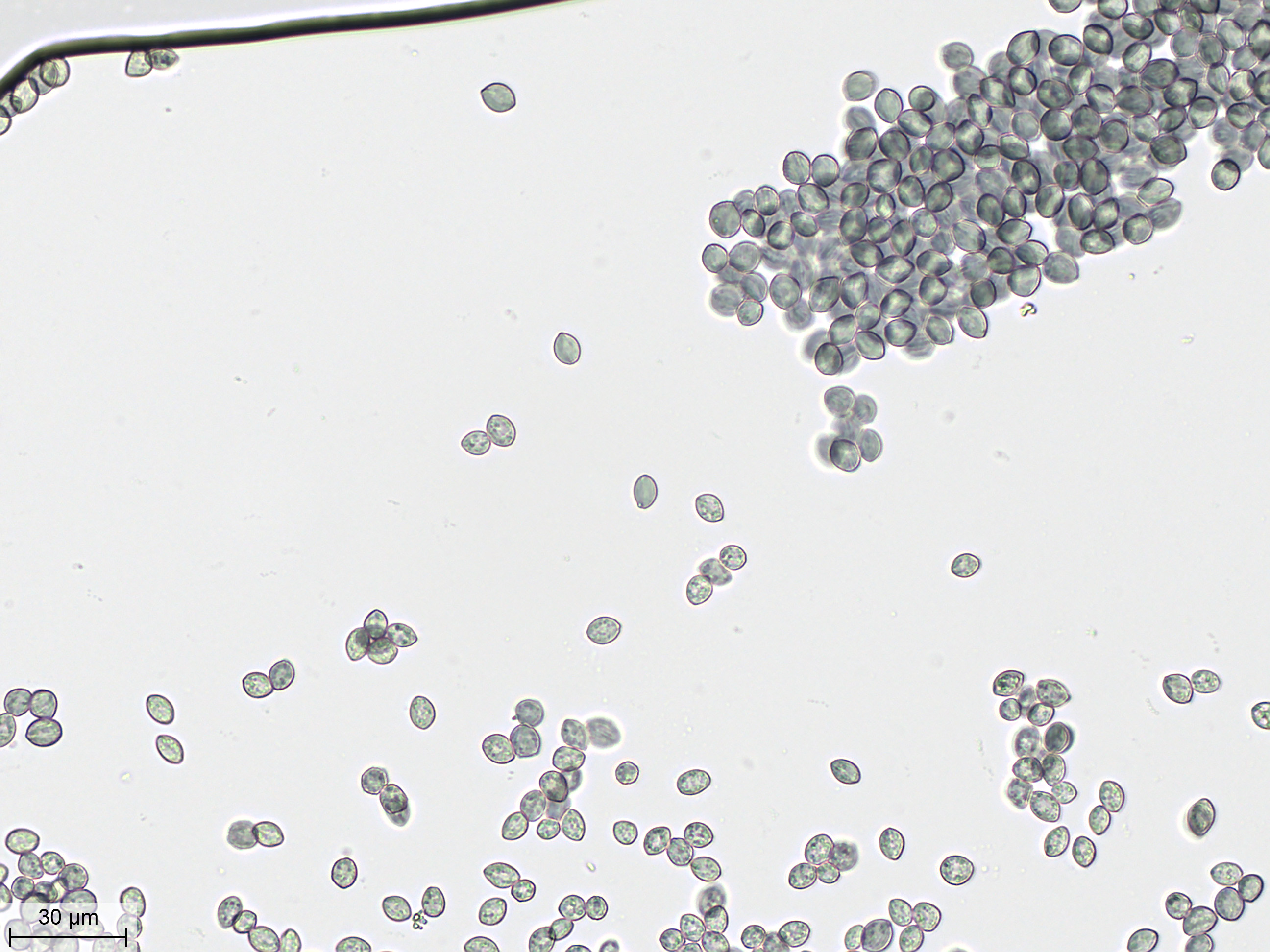 |
||
| Une columelle à l'objectif 40X. Sa membrane est froissée. Elle contient une centaine de spores au centre, mais c'est surtout à l'extérieur que des milliers de spores s'agglutinent. | ||
 |
||
| Et voila, j'ai terminé ! C'était une observation inédite qui m'a encore beaucoup intéressé. Il faudra que j'essaie de faire pousser d'autres sortes de moisissures en changeant de substrat. |